Chaque jour, vous portez une cuillère à votre bouche et instantanément, votre cerveau décode une symphonie de sensations complexes. Cette expérience gustative, apparemment si simple, mobilise en réalité un arsenal sophistiqué de récepteurs sensoriels, de voies neurologiques et de processus biochimiques. Comprendre les mécanismes qui orchestrent cette perception révèle la fascinante complexité de notre système gustatif et explique pourquoi certaines saveurs nous transportent tandis que d’autres nous rebutent. La science moderne dévoile aujourd’hui les secrets de cette machinerie biologique remarquable qui transforme les molécules chimiques en plaisir culinaire.
Anatomie fonctionnelle de la cavité buccale et récepteurs sensoriels
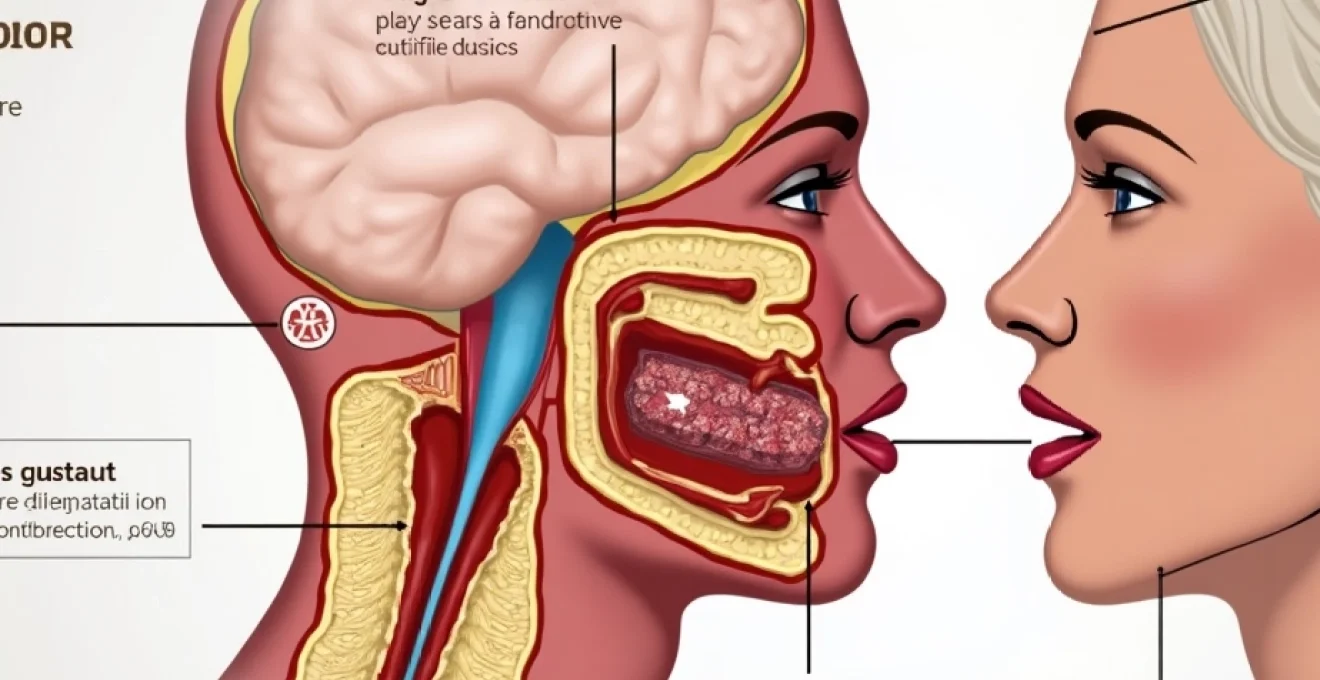
La bouche constitue un véritable laboratoire sensoriel où s’orchestrent les premières étapes de la perception gustative. Cette cavité complexe abrite différents types de structures spécialisées, chacune jouant un rôle précis dans la détection et la transmission des informations gustatives vers le cerveau.
Papilles gustatives : caliciformes, fongiformes et filiformes
Les papilles gustatives se répartissent selon trois morphologies distinctes sur la surface linguale. Les papilles caliciformes , les plus volumineuses, forment un V inversé à l’arrière de la langue et concentrent le plus grand nombre de bourgeons gustatifs. Leur structure en coupe leur permet de retenir efficacement les substances sapides dissoutes dans la salive.
Les papilles fongiformes , reconnaissables à leur forme de champignon, parsèment la pointe et les bords de la langue. Bien que moins nombreuses que les filiformes, elles jouent un rôle crucial dans la détection des saveurs sucrées et salées. Leur couleur rougeâtre, due à leur vascularisation importante, les rend facilement identifiables à l’œil nu.
Les papilles filiformes , les plus abondantes, ne contiennent pas de bourgeons gustatifs mais participent à la texture des aliments grâce à leur surface rugueuse. Elles facilitent la manipulation du bol alimentaire et contribuent aux sensations tactiles pendant la mastication.
Bourgeons gustatifs et cellules de type I, II et III
Chaque bourgeon gustatif constitue une structure ovoïde d’environ 50 à 100 cellules spécialisées. Ces micro-organes sensoriels renferment trois types cellulaires distincts aux fonctions complémentaires. Les cellules de type I , les plus nombreuses, assurent principalement un rôle de soutien structural et de régulation ionique.
Les cellules de type II représentent les véritables détecteurs chimiques des saveurs sucrée, amère et umami. Elles expriment les récepteurs gustatifs couplés aux protéines G et traduisent la liaison des molécules sapides en signaux électriques. Ces cellules libèrent l’ATP comme neurotransmetteur principal vers les fibres nerveuses afférentes.
Les cellules de type III , moins nombreuses mais tout aussi essentielles, se spécialisent dans la détection de l’acidité et du goût salé. Elles établissent des synapses conventionnelles avec les neurones sensoriels et libèrent de la sérotonine et du GABA. Leur morphologie bipolaire facilite la transmission rapide des signaux gustatifs.
Innervation cranienne : nerfs facial, glossopharyngien et vague
L’innervation de la cavité buccale fait appel à trois nerfs crâniens majeurs qui acheminent les informations gustatives vers le système nerveux central. Le nerf facial (VII) innerve les deux tiers antérieurs de la langue via sa branche corde du tympan, transmettant principalement les sensations sucrées et salées.
Le nerf glossopharyngien (IX) prend en charge le tiers postérieur de la langue, région particulièrement sensible aux saveurs amères. Cette innervation stratégique constitue un mécanisme de protection contre l’ingestion de substances potentiellement toxiques. Le nerf vague (X) complète cette innervation en desservant l’épiglotte et certaines zones pharyngées.
Glandes salivaires submandibulaires et parotidiennes
La production salivaire, orchestrée par les glandes submandibulaires, parotidiennes et sublinguales, conditionne directement la perception gustative. Les glandes submandibulaires produisent une salive mixte, riche en mucines et en enzymes, qui facilite la dissolution des molécules sapides et leur contact avec les récepteurs.
Les glandes parotidiennes sécrètent une salive séreuse, pauvre en mucus mais riche en amylase et en protéines antimicrobiennes. Cette composition particulière influence la perception des saveurs en modifiant l’environnement chimique buccal. La composition individuelle de la salive explique en partie pourquoi vous percevez certains goûts différemment de vos proches.
Mécanismes neurobiologiques de la transduction gustative
La transformation des stimuli chimiques en signaux électriques constitue l’essence même de la perception gustative. Ce processus, appelé transduction sensorielle, fait intervenir des mécanismes moléculaires spécifiques pour chaque saveur fondamentale. La compréhension de ces voies de signalisation révèle la sophistication remarquable de notre système gustatif.
Canaux ioniques ENaC pour la perception du salé
La détection du sodium fait principalement appel aux canaux ioniques ENaC (Epithelial Sodium Channels) exprimés dans les cellules gustatives de type III. Ces canaux permettent l’entrée directe des ions sodium dans la cellule, provoquant une dépolarisation membranaire proportionnelle à la concentration saline.
Cette perméabilité sélective au sodium explique pourquoi le chlorure de sodium génère une sensation salée pure, contrairement au chlorure de potassium qui présente un arrière-goût amer. L’activation des canaux ENaC déclenche l’ouverture de canaux calciques voltage-dépendants, amplifiant le signal et déclenchant la libération de neurotransmetteurs.
Récepteurs T1R2-T1R3 et détection des molécules sucrées
La reconnaissance des substances sucrées repose sur l’hétérodimère T1R2-T1R3 , un récepteur métabotrope couplé aux protéines G. Cette combinaison unique de sous-unités permet la détection d’une remarquable diversité de molécules sucrées, du saccharose aux édulcorants artificiels en passant par les acides aminés comme la glycine.
L’activation de ce récepteur déclenche une cascade de signalisation impliquant l’adénylyl cyclase, l’élévation de l’AMPc intracellulaire et finalement la libération d’ATP. Cette voie de transduction explique la sensibilité extrême aux substances sucrées et la capacité à détecter des concentrations infimes d’édulcorants puissants comme le stéviose.
Récepteurs T2R et reconnaissance de l’amertume
L’amertume fait intervenir une famille de 25 récepteurs T2R chez l’humain, chacun spécialisé dans la détection de classes particulières de composés amers. Cette diversité réceptorielle témoigne de l’importance évolutive de la détection des toxines végétales, nombreuses et chimiquement variées.
Contrairement aux autres saveurs, l’amertume présente une architecture de détection redondante où plusieurs récepteurs peuvent reconnaître la même molécule. Cette organisation assure une sensibilité maximale aux composés potentiellement dangereux. L’activation des récepteurs T2R suit la même voie de signalisation que les récepteurs sucrés, aboutissant à la libération d’ATP.
Canaux PKD2L1 et transduction de l’acidité
La perception de l’acidité implique principalement les canaux PKD2L1 (polycystic kidney disease 2-like 1), sensibles aux variations de pH. Ces canaux permettent l’entrée d’ions calcium en réponse à l’acidification du milieu extracellulaire, déclenchant directement la dépolarisation des cellules gustatives de type III.
La transduction acide présente la particularité d’être directement proportionnelle à la concentration en protons libres, expliquant pourquoi vous percevez l’intensité de l’acidité de manière logarithmique. Cette relation permet de discriminer efficacement les différents niveaux d’acidité rencontrés dans l’alimentation.
Umami et activation des récepteurs mGluR4
La saveur umami, reconnaissance du glutamate et de l’aspartate, fait appel à l’hétérodimère T1R1-T1R3 mais également aux récepteurs mGluR4 (récepteurs métabotropiques au glutamate de type 4). Cette double voie de détection confère à l’umami ses caractéristiques particulières, notamment les phénomènes de synergie avec les nucléotides.
L’interaction entre l’inosine monophosphate (IMP) et le glutamate amplifie considérablement la perception umami, expliquant l’efficacité des exhausteurs de goût utilisés en cuisine asiatique. Cette synergie révèle la complexité des interactions moléculaires dans la perception gustative.
Processus de mastication et libération aromatique
La mastication déclenche une cascade de phénomènes physiologiques qui transforment radicalement l’expérience sensorielle. Ce processus mécanique ne se contente pas de broyer les aliments ; il libère les molécules volatiles emprisonnées dans la matrice alimentaire et facilite leur migration vers les récepteurs olfactifs par la voie rétronasale.
La fragmentation des structures cellulaires végétales ou animales pendant la mastication expose de nouvelles surfaces aux enzymes salivaires. L’amylase salivaire commence la dégradation des amidons, libérant des sucres simples qui modifient progressivement la perception sucrée. Simultanément, la lipase linguale attaque les triglycérides, générant des acides gras libres responsables des sensations de gras.
Les mouvements de la langue durant la mastication créent des flux d’air qui propulsent les composés volatiles vers la cavité nasale. Cette ventilation rétronasale explique pourquoi la texture des aliments influence leur perception aromatique. Les aliments croustillants génèrent des flux d’air plus importants, intensifiant la libération aromatique.
La température de l’aliment module considérablement la volatilité des composés aromatiques et donc leur perception olfactive.
La formation du bol alimentaire implique une coordination précise entre la sécrétion salivaire et les mouvements masticatoires. La salive agit comme un solvant universel, permettant la dissolution des molécules sapides et leur transport vers les récepteurs gustatifs. Sa composition enzymatique transforme également certains précurseurs inertes en molécules actives sensoriellement.
Voies neurologiques centrales du goût
L’information gustative suit un parcours neurologique complexe depuis les récepteurs périphériques jusqu’aux centres corticaux supérieurs. Cette organisation hiérarchique permet une intégration progressive des signaux sensoriels et leur transformation en perception consciente. Chaque relais neurologique apporte sa contribution spécifique au traitement de l’information gustative.
Noyau du tractus solitaire et intégration primaire
Le noyau du tractus solitaire (NTS) constitue la première station d’intégration centrale des signaux gustatifs. Cette structure médullaire reçoit les afférences des nerfs crâniens VII, IX et X et organise l’information selon une topographie rostro-caudale qui respecte partiellement l’organisation des saveurs.
Les neurones du NTS présentent déjà une spécialisation fonctionnelle, certains répondant préférentiellement aux substances sucrées, d’autres aux composés amers ou salés. Cette spécialisation précoce suggère que le codage des saveurs commence dès les premières étapes du traitement central. L’intégration à ce niveau permet également les premiers mécanismes de plasticité gustative.
Thalamus ventral postérieur médian et relais sensoriel
Le thalamus ventral postérieur médian (VPM) constitue le relais thalamique obligatoire vers le cortex gustatif. Cette structure sous-corticale assure un filtrage sophistiqué de l’information gustative, modulant l’intensité des signaux en fonction du contexte physiologique et comportemental.
Les neurones thalamiques reçoivent également des afférences du système limbique, permettant une modulation émotionnelle précoce de la perception gustative. Cette convergence explique pourquoi votre état émotionnel influence directement la perception des saveurs. Le VPM joue également un rôle crucial dans l’attention gustative sélective.
Cortex gustatif primaire insulaire antérieur
Le cortex insulaire antérieur représente l’aire corticale primaire de traitement de l’information gustative. Cette région présente une organisation en colonnes où chaque colonne traite préférentiellement une saveur particulière. L’activité neuronale de cette aire corrèle directement avec l’intensité perçue des stimuli gustatifs.
L’insula antérieure intègre également les informations trigéminales liées à la texture, à la température et aux sensations irritantes. Cette convergence multimodale constitue les bases neurobiologiques de la perception de flaveur. Les lésions de cette région provoquent des troubles gustatifs sévères, confirmant son rôle central.
Cortex orbitofrontal et perception hédonique
Le cortex orbitofrontal transforme l’information gustative brute en expérience hédonique consciente. Cette région évalue la valeur récompensante des aliments en intégrant les signaux gustatifs, olfactifs, visuels et contextuels. L’activité orbitofrontale détermine si vous trouvez un aliment plaisant ou déplaisant.
Cette aire corticale présente une plasticité remarquable,